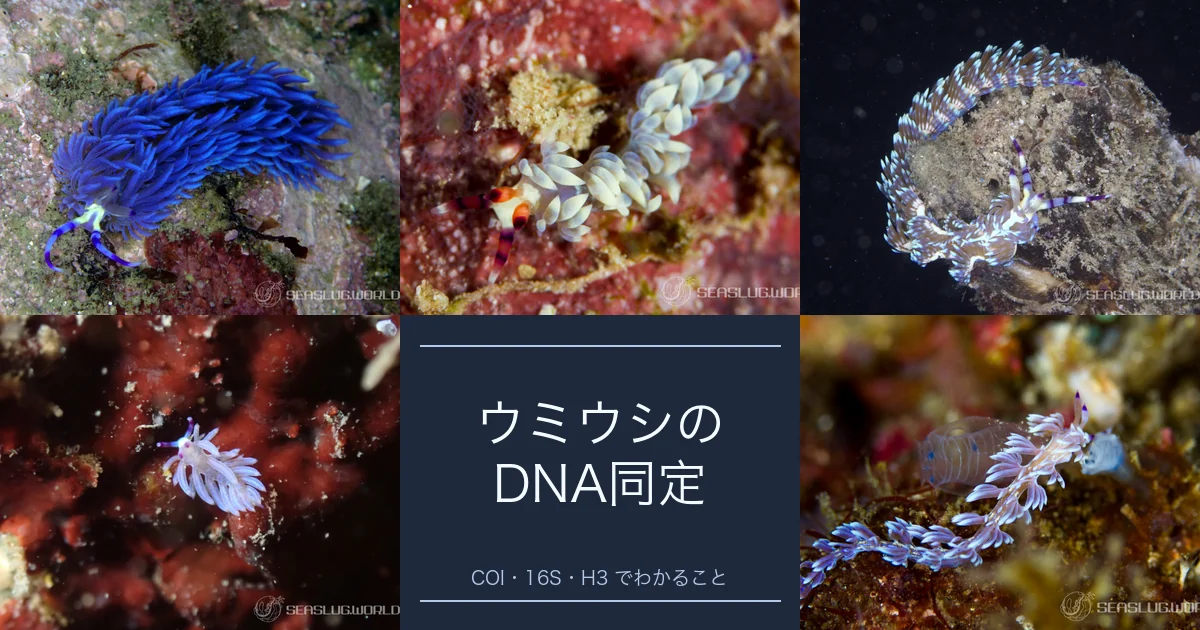
スミゾメキヌハダウミウシは 4 種の複合だった:馬場の 4 mm 1 個体から 65 年
ハゼの鰭にしがみついて、 鰭条のあいだの薄い皮膚を齧り取る黒いウミウシ — スミゾメキヌハダウミウシ Gymnodoris nigricolor。 ダイバーに長く知られてきたこの種が、 De Souza-Canal et al. (2025) で 少なくとも 4 種からなる種複合体 に分解されました。 元になっているのは、 馬場菊太郎が 1960 年に三崎・相模湾の泥底から拾った 体長 4 mm、 1 個体のみ の標本。 そこから 65 年経って、 ようやく「あの黒いウミウシ」 の内訳が見えはじめたところです。
論文
馬場 1960 の原記載 — 三崎の 4 mm 1 個体
Gymnodoris nigricolor Baba, 1960 はそもそも、 神奈川県三浦半島の三崎、 相模湾内の水深 2 m の泥底から採集された たった 1 個体・体長 4 mm から記載された種です。
原記載で馬場が書いた特徴はおおまかにこう。
種小名 nigricolor はラテン語 niger (黒) + color (色)。 和名「スミゾメ (墨染)」 も同じく全身の黒色を表していて、 種小名と意味が対応します。
以後 65 年、 三崎・相模湾からこの種の追加標本がきちんと記録された形跡はありません。 つまり「Gymnodoris nigricolor」 の参照点は、 ずっと馬場 1960 のこの 1 枚の図と 4 mm の 1 個体だった、 ということです。
「ハゼの鰭を齧るウミウシ」 という有名な顔
スミゾメキヌハダウミウシがダイバーに知られているのは、 圧倒的に ハゼの鰭への異常な執着 のせいです。
- Mulliner 1991 — ダテハゼ類の鰭にしがみついた個体を初記録
- Osumi & Yamasu 1994, 2000 — 鰭条のあいだの薄い皮膚を齧り取って摂食する行動と、 初期発生を記載
ハゼ自身は食べ尽くされず、 1 尾のハゼに複数のスミゾメが同時に取り付くこともある。 寄生に近い片利、 という整理がされてきました。 同属のほかの種 (キヌハダウミウシ G. inornata や オキナワキヌハダウミウシ G. okinawae など) はおもに別のウミウシやその卵を食べているので、 「ハゼに付くウミウシ」 は実質スミゾメ 1 種だけ、 という認識が長く続いてきました。
報告地はインド洋・紅海まで広がり (Bidgrain 2014)、 「全身一様な黒のウミウシなんてほかにいない、 だから世界中の黒いキヌハダはぜんぶこれ」 という運用がされてきた、 というのが分解前の状況です。
ただし伏線はあって、 Knutson & Gosliner (2022) がマーシャル諸島・クワジェリン環礁の「G. cf. nigricolor」 を配列だけ NCBI に登録した時点で、 これが本物の nigricolor から系統的にかなり離れていることはすでに見えていました。
De Souza-Canal, Nakano & Valdés (2025) の解像度
今回の論文は ニューカレドニア (Koumac、 Poum) と 沖縄・奄美 から計 14 個体を集め、 COI + 16S rRNA + Histone H3 の 3 遺伝子で系統解析し、 ASAP と bPTP の 2 種の種境界推定法で種数を割り出しています。
結論はシンプル。 G. nigricolor は 少なくとも 4 系統 に分かれ、 そのうち 2 つは新種として記載され、 1 つは別研究者が記載作業中、 1 つは三崎の原記載に同定される、 という整理です。
4 種の内訳
Gymnodoris nigricolor (狭義) — 馬場 1960 の原記載が指す種

分布は三崎 (タイプ産地) と ニューカレドニア・クマック。 識別の決め手は 触角の基部が左右で融合していること。 歯舌の内側歯は短く、 中央の鋭い歯尖の両側に平らな歯尖が並ぶ三尖型で、 これも馬場 1960 の図と一致します。
ただし、 著者らは 三崎のタイプ産地から新規標本を入手できていません。 「Koumac 個体が馬場 1960 の図と形態的に一致する」 という間接的な根拠で、 ニューカレドニア標本を狭義 (sensu stricto) と判定している、 という整理。 つまり G. nigricolor の同定の基準そのものが、 いま ニューカレドニア標本 に乗っかっています。 三崎でのタイプ産地からの新規採集と DNA 照合は、 今後の課題として残っています。
Gymnodoris boussionae De Souza-Canal, Nakano & Valdés, 2025 — ニューカレドニア固有
ニューカレドニアの Koumac と Poum から 10 個体。 一見すると G. nigricolor と区別がつかないベルベットの黒ですが、 触角の基部は分離していて融合しない。 内側の歯は強く湾曲して細長く、 鋸歯はなし。 ペニスの棘も短く湾曲、 前立腺は扁平で幅広。
種小名 boussionae は、 採集に協力したニューカレドニアのダイバー兼水中ビデオグラファー Magali Boussion への献名。 摂食行動は観察されていません。
Gymnodoris nagoensis De Souza-Canal, Nakano & Valdés, 2025 — 沖縄・奄美

ホロタイプは沖縄県名護市 (10 m)、 他に沖縄県宜野湾市の砂辺と鹿児島県奄美大島の手広海岸。 種小名 nagoensis は名護市にちなんでいます。
外見的には触角が他種より細長く、 基部は分離。 鰓は 10 枚の単純な鰓葉のみで、 ほかの 3 種より少なめ。 著者らは G. nagoensis の個体が ハゼの巣穴に這い込んで鰭を齧る行動を観察 していて、 1 尾のハゼに複数個体が同時に取り付くことも記録しています。 つまり 4 種の中でハゼ鰭の摂食が現場で直接確認されているのは、 今のところ G. nagoensis だけ、 ということになります。
なお、 系統樹上では G. nagoensis は G. nigricolor (狭義) の姉妹群で、 統計的支持もきわめて強く、 遺伝的にはかなり近い間柄。 歯舌は G. boussionae によく似ているけれど、 これは食性収斂による可能性が示唆されています。
マーシャル諸島の未記載種
Knutson & Gosliner (2022) が G. cf. nigricolor として配列登録したクワジェリン環礁の個体。 系統上は上 3 種からはっきり離れます。 形態記載は他研究グループが別論文で進めているとのこと (Gosliner 私信)。 さらに Mehrotra et al. (2021) が タイ・コタオから「鰓が白い」 個体を示しており、 著者らはこれも別種候補と書いています。 つまり「4 種」 は最低ラインで、 紅海・タイ・フィリピン側にはまだ未記載の系統が残っている見通しです。
なお、 同じ著者ら (Nakano 抜きの De Souza-Canal & Valdés) は、 本論文の 4 か月後にあたる 2025 年 10 月、 Zootaxa にニューカレドニア産 Gymnodoris 属の 73 ページ monograph を出しており、 そこで knutsonae / fleuroti / mariefranceae / lebouteillerorum / deniseae / bricei / wanaporum / fassioae / feyae / bernardae / boiteuxi の 11 新種 を一気に記載しています (Souza-Canal & Valdés 2025, Zootaxa 5710(1):1-73)。 ニューカレドニアだけで本属の biodiversity がここまで膨らんだ、 という背景もあわせて押さえておくと、 Knutson & Gosliner (2022) が genus 全体で「60-70 候補種、 大半は未記載」 と書いた状況がいま急速に消化されつつあることが見えてきます。
ダイバー目線で何が変わるか
「ハゼに付いてる黒いウミウシ = スミゾメ」 は崩れた
過去 30 年でハゼ寄生として記録されてきた個体のうち、 南日本での観察は 今後 G. nagoensis の可能性 が出てきました。 著者ら自身、 「4 種のうち nagoensis 以外の摂食行動は観察できていない」 「すべての種が必ずしもハゼの鰭を食べているとは限らない」 と明記しています。 つまり「ハゼ寄生 = G. nigricolor」 という古い構図は、 文献の同定帰属まで遡って書き換わる余地があります。
でも、 撮影されたハゼ採食シーンからは同定できない
ここが今回いちばん効いてくる現実です。 4 種を分ける外見の決め手は 触角の基部の融合・分離 で、 G. nigricolor (狭義) のみ融合、 G. boussionae と G. nagoensis は分離。 鰓の枚数 (nigricolor 10〜11 / boussionae 16〜19 / nagoensis 10) も補助的に効きます。
ところが、 ダイバーが撮るハゼ採食シーンの写真は、 だいたい次のどちらかです。
- ハゼの鰭にしがみついている個体の全身ショット
- 体の側面 / 斜め上アップ

触角の基部 (左右が引っ付いているかどうか) は、 ハゼに張り付いた姿勢だと頭がハゼの鰭側に向いていることが多く、 真上からのアングルにならない限り写りません。 鰓の枚数もハゼに巻き付いた体勢ではほぼ数えられません。 つまり 「ハゼに付いてる黒いウミウシ」 だけでは 4 種のどれかを決められない のが、 写真ベースの観察記録としての結論になります。
「あー、 過去のあの写真、 同定書き換えしたいけど、 そもそも書き換えるための情報が画面に写ってない」 — というのが、 こちらの率直な感触です。
三崎産の再採集が要る
タイプ産地の三崎 (神奈川県三浦半島) から、 馬場 1960 のあとに新規標本が記録されていません。 今回の G. nigricolor (狭義) の同定根拠は、 ニューカレドニア標本が馬場 1960 の図と形態的に一致するから、 という間接認定。 将来、 三崎で新規個体が採れて DNA が回収できれば、 場合によっては「三崎産は今回 狭義とされた系統と別物」 という整理に変わる可能性も残ります (タイプ標本の保存状態によっては DNA が取れない、 ということも当然ありえます)。
三崎の泥底 2 m はダイバーアクセス可能なエリアでもあるので、 ハゼ採食でない単独個体を見たら、 ぜひ触角の基部と鰓を真上から押さえた 1 枚を残しておいてください。
サイトへの反映 (済)
- スミゾメキヌハダウミウシ Gymnodoris nigricolor の種ページに、 4 種分解後の整理を反映済
- Gymnodoris boussionae (ニューカレドニア固有) を新規ページとして登録済
- Gymnodoris nagoensis (沖縄・奄美産) を新規ページとして登録済
過去の沖縄・奄美のハゼ採食観察記録写真については、 今後 G. nagoensis 側に紐付け直す方向で整理を検討しています。 ただし上に書いたとおり、 採食シーン写真は触角基部が写っていないことが多く、 全部を機械的に書き換えることはできません。 触角基部が写っている個体のみ移し替えていく予定です。
おわり
馬場 1960 の 4 mm 1 個体から 65 年。 「ハゼに付いてる黒いウミウシ」 という素朴な印象論で長くまとめてきた対象が、 最低 4 系統の総称だった、 という整理に切り替わります。 次に動くのは三崎タイプ産地からの再採集と、 タイ・紅海側の未記載系統の記載のはず。
撮影では 触角の基部 と 鰓 を真上から押さえた 1 枚を、 ハゼ採食シーンとは別に残しておくと、 後から同定し直せる材料になります。 単独で這っている個体に出会ったら、 ぜひ 写真投稿フォーム から登録してください。
この記事が良かったら、 著者にチップを送れます。
この記事をチップで応援